|
O
estudo do sono tem despertado interesse desde a antiguidade, suscitando
diversos documentos escritos a cerca deste assunto. Segundo Timo-Iaria
(1985), o sono foi objeto de análise subjetiva, observativa e, sobretudo
discursiva durante milhares de anos em todas as civilizações.
Até o
presente momento, a função do sono ainda não está completamente
esclarecida, mas, sabe-se que serve para restaurar os níveis normais de
atividade e o “equilíbrio” normal entre as diferentes partes do sistema
nervoso central (Guyton e cols, 1997), além de estar envolvido com a
conservação do metabolismo energético, com a cognição, com a
termorregulação, com a maturação neural e a saúde mental (Kandel e cols,
2003).
Embora represente um período de descanso, o sono parece não ser um
indicador de estado passivo do sistema nervoso, correspondendo então a
um fenômeno ativo, com fases distintas e bem caracterizadas (Aserinsky e
Kleitman, 1953; Crick e Mitchison, 1983).
Considerando o aspecto científico, foi a partir da década de 30 que
avanços notáveis a cerca do estudo do sono possibilitaram um maior
progresso e compreensão do assunto.
Berger (1930) obteve um grande avanço no estudo dos mecanismos de sono e
vigília quando registrou a atividade elétrica encefálica, por meio de
eletrodos de agulhas introduzidos no couro cabeludo (eletroencefalograma,
ou EEG). Nos primeiros EEGs registrados durante o sono, Berger descreveu
os fusos e as ondas delta, que caracterizam as fases II, III e IV do
sono humano, descritas mais adiante.
O
sono apresenta estágios alternantes, e cada estágio possui padrões
eletroencefalográficos característicos, segundo freqüência e amplitude
das ondas, que são distintos daqueles observados durante a vigília (Loomis
e cols., 1937; 1938).
Rimbaud e cols. (1955) observaram a ocorrência de curtas fases
dessincronizadas no EEG durante o sono natural de gatos, que eram
semelhantes ao padrão típico de atividade elétrica no hipocampo (ritmo
teta). Essas fases constituem um estado de sono inteiramente distinto
daquele com padrão sincronizado e foram denominadas de “sono
dessincronizado” (Moruzzi, 1972) ou também “sono paradoxal” (Jouvet,
1972), pois possui padrão eletroencefalográfico análogo ao da vigília,
porém associado à ausência do tônus muscular (Jouvet e Michel, 1959). O
sono paradoxal passou a ser extensamente estudado, em especial devido
aos movimentos periódicos rápidos dos olhos na espécie humana (por esse
motivo, esse estágio também pode ser denominado sono REM – “Rapid Eye
Movements”).
Esta
fase também se caracteriza pela presença de sonhos (Dement e Kleitman;
1957), pelos eventos fásicos (que incluem os movimentos oculares
rápidos, e também abalos musculares ou mioclonias, e ondas
ponto-genículo-occipitais - PGO) e tônicos (as já citadas atonia
muscular esquelética e dessincronização do eletroencefalograma cortical,
ou seja, ondas de baixa amplitude e alta freqüência, e flutuações
cardio-respiratórias) (Aserinsky e Kleitman, 1953; Jouvet, 1962, Vertes,
1984; Siegel, 1994).
Estudos demonstram que o tronco encefálico, particularmente a formação
reticular pontina lateral e bulbar medial, é a área responsável pela
geração do sono REM, conforme revisado por Vertes (1984) e Siegel
(1994).
Em
contrapartida ao sono REM, existe o sono NREM (“Non-Rapid Eye Movements”),
que também pode ser denominado sono sincronizado, pois um potencial
elétrico rítmico inibitório-excitatório, gerado por neurônios talâmicos
e corticais, forma ondas sincronizadas de alta amplitude e baixa
freqüência (Steriade, 1992). Esta fase do sono pode ainda ser
subdividida em quatro estágios conforme o aumento de sua profundidade (Rechtschaffen
e Kales, 1968):
Estágio 1: o indivíduo se encontra numa transição entre o estado
de vigília e o sono, com ondas no EEG de baixa voltagem;
Estágio 2: presença de ondas no EEG de baixa voltagem, sendo
interrompido por fusos e complexo K, que são ondas de alta amplitude;
Estágio 3: presença de ondas delta, que possuem baixa
freqüência e alta amplitude;
Estágio 4: predominância de ondas delta, lentas. Juntamente com o
estágio 3, é denominado também como ‘Sono de Ondas Lentas’.
De modo geral, o sono NREM apresenta baixa atividade neuronal, taxa
metabólica baixa, temperatura encefálica baixa, declínio da atividade
simpática e aumento da atividade parassimpática, pouca atividade
muscular e a regulação da temperatura está presente (Kandel e cols,
2003).
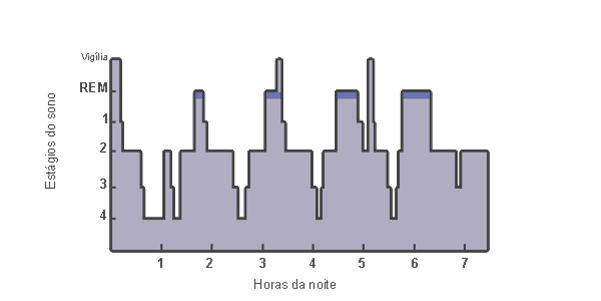
Durante o período de sono em seres humanos normalmente ocorrem de 4 a 6
ciclos
bifásicos (Figura 1) com
duração de 90 a 100 minutos cada, sendo cada um dos ciclos composto
pelas fases do sono NREM, com duração de 45 a 84 minutos, e pela fase do
sono REM, que dura de 5 a 45 minutos (Horne, 1980; Morrison, 1983).
Em indivíduos adultos normais, o sono NREM encontra-se principalmente
distribuído na primeira metade do período de sono, enquanto o sono REM
predomina na segunda metade deste período. Em seres humanos, um adulto
jovem saudável, com ciclo vigília-sono convencional,
apresenta, aproximadamente, as seguintes proporções em relação ao tempo
total de sono (Carskadon e Dement, 1994):
estágio 1 → 2 - 5%,
estágio 2 → 45 - 55%,
estágio 3 → 8%,
estágio 4 → 10 - 15%,
Figura
1:
O ciclo dos estágios do sono de seres humanos de vida adulta inicial.
Pode-se observar que o sono caracteriza-se pela progressão do estágio 1
para o estágio 4 do sono NREM. Depois de aproximadamente 70-80 minutos,
o individuo retorna brevemente para os estágios 3 ou 2, e então entra na
primeira fase do sono REM da noite, que dura aproximadamente 5-10
minutos. Nos seres humanos, a duração do ciclo representado pelo início
do sono NREM até o final da primeira fase de sono REM é de
aproximadamente 90-110 minutos. Este ciclo sono NREM/REM é repetido de 4
a 6 vezes durante a noite. Nos ciclos sucessivos a duração dos estágios
3 e 4 do sono NREM diminuem à medida que a duração da fase REM aumenta.
Fonte: Kandel e cols, 2001.
O ritmo circadiano básico do
ciclo sono-vigília é gerado pelo núcleo supraquiasmático, geralmente
vinculado às oscilações na temperatura e ao ciclo claro-escuro geofísico
(Moore-Ede e cols., 1982). Em seres humanos normalmente, o período de
sono ocorre durante a noite e geralmente é único. Em ratos e
camundongos, por exemplo, o período de sono é durante o dia, e o sono é
fragmentado, característica de animal predado.
Sagüis (Callithrix sp.) podem ser bons modelos de sono para o
sono humano, pois possuem o período de sono durante a noite. Além disso,
estes animais apresentam as mesmas fases (NREM e REM) e os mesmos
estágios do sono NREM, em forma de ciclos bifásicos tal qual o humano,
demonstrado por
Crofts
e colaboradores (2001), ao serem analisados por radiotelemetria.
Utilizando essa técnica (telemetria), além do registro do
eletroencefalograma, informações sobre a temperatura corpórea e a
atividade locomotora também podem ser obtidas. Não obstante, a restrição
física ao animal é inexistente, ao contrário do que aconteceu em
experimentos que fixavam o animal em uma cadeira (Weitzmann, 1961), ou
acoplavam ao animal uma mochila contendo a aparelhagem para medição (Bert
e cols, 1975).
Utilizando-se a radiotelemetria, portanto, não há (ou pelo menos é muito
reduzida quando comparada ao registro convencional) restrição física e
psicológica ao animal, e este pode continuar vivendo com seu grupo, no
local onde já está acostumado, possibilitando, desta maneira, uma maior
confiabilidade do resultado.
Até o momento, poucos estudos utilizando a radiotelemetria foram feitos
com o objetivo de analisar o sono em sagüis. O trabalho desenvolvido
neste laboratório baseia-se em estudos de transmissores implantados no
encéfalo de sagüis, possibilitando o registro de um canal de EEG durante
o período de sono. Os diferentes estágios do sono são classificados de
acordo com os critérios de Rechtschaffen e Kales (1968), exceto pela
ausência das informações do eletrooculograma e eletromiograma, em
vigília, provável REM (pREM), sono leve (estágios 1 e 2) e sono de ondas
lentas (estágios 3 e 4).
Para melhor caracterizar os estágios do sono, informações adicionais,
como registro do eletromiograma e da temperatura, podem ser obtidas.
Além disso, o trabalho de Crofts e seus colaboradores abre
possibilidades para o estudo de drogas que modulam o sono, neste modelo
comprovadamente adequado.
Assim, a
proposta do presente trabalho é analisar a resposta eletrofisiológica e
comportamental do sono de sagüis às seguintes drogas: modafinil (droga
que age diminuindo o sono), diazepam (droga indutora de sono) e
donepezil (droga que aumenta a frequência do sono REM). Essas drogas são
utilizadas na clínica no tratamento de narcolepsia (Bastuji e cols,
1988; Boivin e cols, 1993), para o tratamento de insônia (Gilman, 1996),
e no tratamento da Doença de Alzheimer (Geldmacher, 1997),
respectivamente.
Obs.:
Clique aqui para mais informações
|
Rerências Bibliográficas: |
Aserinsky, E; Kleitman, N – Regularly occurring periods of eye motility
and concomitant phenomena during sleep. Science, 118:273-274,
1953.
Bastuji, H; Jouvet, M – Successful treatment of idiopathic hypersomnia
and narcolepsy with Modafinil. Prog Neuroosychopharmacol Biol
Psychiatry. 12:695-700, 1988.
Berger, H – Ueber das elektrenkephalogramn des menschen. J. Physiol.
Neurol., 40:160-179, 1930.
Bert, J; Balzamo, E; Chase, M; Pegram, V – The sleep of the Baboon,
Papio papio, under natural conditions and in the laboratory.
Electroenceph. Clin. Neurophysiol; 39:657-662,
1975.
Boivin, DB; Montplaisir, J; Petit, D; Lambert, C; Lubin, S – Effects of
Modafinil on the symptomatology os human narcolepsy. Clin
Neuropharmacol. 16(1):46-53, 1993
Carskadon, MA; Dement, WC – Normal human sleep: an overview. Em Kryger,
MH; Roth, T; Dement, WL; Eds – Principles and practice of sleep medicine.
1° ed. Philadelphia, W. B. Sounders Company. p. 943-960, 1989.
Crick, F; Mitchison, G – The function of dream sleep. Nature,
304:111-114, 1983.
Crofts, HS; Wilson, S; Muggleton, NG; Nutt, DJ; Scott, EAM; Pearce, PC
– Investigation of the sleep electrocorticogram of the common marmoset (Callithrix
jacchus) using radiotelemetry.
Clinical Neurophysiology; 112:2265-2273, 2001.
Dement, W; Kleitman, N – Cyclic variations in EEG during sleep and
their relation to eye movements, body motility, and dreaming.
Electroencephalogr. Clin. Neurophysiol.,
9:673-690, 1957.
Gilman, AG; Hardman, JG; Limbird, LE; Molinoff, PB; Ruddon, RW – As
bases farmacológicas da terapeutica; Editora Mc Graw Hill; nona edição,
160-162, 1996.
Guyton, AC; Hall, JE – Tratado de fisiologia médica; Editora Guanabara
Koogan; nona edição, 685-689, 1997.
Horne,
JA – Sleep and body restitution. Experientia. 36:11-13,
1980.
Jouvet,
M; Michel, F – Corrêlations électromyographique do sommeil chez lê chat
décortiqué et mésencéfalique chronique. Compt. Rend. Soc. Biol.,
153:422-425, 1959.
Jouvet,
M – Récherces sur les structures nerveuses et les mecanismes resonsables
des differentes phases du sommeil physiologyque.
Arch. Ital. Biol.,
100:125-296, 1962.
Jouvet,
M – The role of monoamines and acetylcholine-containg neurons in the
regulation of the sleep-waking cycle. Ergedhisse der Physiologie.
64:166-307, 1972.
Kandel,
ER; Schwartz, JH; Jessel, TM – Princípios da neurociência; Editora
Manole; quarta edição, 936-947, 2003.
Loomis,
AL; Harvey, EN; Hobart, GA – Cerebral states during sleep as studied by
human brain potentials. J. Exp. Psychol., 21:127-144,
1937.
Loomis, AL; Harvey, EN; Hobart, GA – Distribution of disturbance
patterns in the human electroencephalogram, with special reference to
sleep . J.Neurophysiol., 1:413-430, 1938.
Moore-Ede, MC; Sulzman, FM; Fuller, CS – Circadian timing of
physiological systems. The clocks that times us. Cambridge, Harvard
University Press. p. 201-294. 1982.
Morrison, AR – A window on the sleeping brain. Sci. Am..
248:94-102, 1983.
Moruzzi, G – The sleep-waking system. Ergedhisse der Physiologie.
64:1-165, 1972.
Pearce, PC; Crofts, HS; Muggleton, NG; Scott, EAM – Concurrent
monitoring of EEG and performance in the common marmoset: a
methodological approach. Physiol Behav. 63:591-599.
Rechtschaffen, A; Kales, A – Manual of standardized terminology,
techniques, and scoring system for sleep stages of human subjects.
Brain Information Service/Brain Research Institute, UCLA, Los
Angeles, 1968.
Rimbaud,
L; Passouant, P; Cadilhac, J – Participacion de l’hippocampe à la
régulation des états de veille et de sommeil. Rev. Neurol., 93:303-308,
1955.
Siegel,
JM – Brainstem mechanisms generating REM sleep. Em: Kryger, MH; Roth, T;
Dement, WC; Ed – Principles and practice of sleep medicine.
2° Ed.
Philadelphia, W. B. Saunders Company. p. 125-144, 1994.
Steriade, M – Basic mechanisms of sleep generation. Neurology,
42 (suppl. 6):9-18, 1992.
Timo-Iaria, C – O sono; Ciência Hoje. 4:66-67, 1985.
Vertes, RP – Brainstem control of the events of REM sleep.
Prog. Neurobiol., 22:241-288, 1984.
Weitzmann, ED; - A note on the EEG and eye movements during behavioral
sleep in monkeys. Electroenceph. Clin. Neurophysiol;
13:790-794, 1961.
(voltar)
|